AtaGenix Laboratories

AtaGenix Laboratories
Release time: 2026-05-08 View volume: 392
Centrosomes must complete maturation during the G2/M phase to activate their microtubule-organizing center function, ensuring proper assembly of the bipolar spindle and accurate chromosome segregation. PLK1 is a key kinase for centrosome maturation, and its spatiotemporal localization at the centrosome directly determines the timing of maturation. However, how the cell cycle regulates the timely recruitment of PLK1 has remained unclear. A recent study published in Nature Communications has, for the first time, revealed that the ubiquitin E3 ligase RNF40 forms a signaling cascade with CDK1-PLK1 through an “acetylation-phosphorylation switch,” thereby precisely controlling the temporal window of centrosome maturation. This discovery not only elucidates the molecular switch mechanism of centrosome maturation but also provides new targets for tumor chromosomal instability and chemotherapy sensitization.
Interaction between RNF40 and PLK1
Using immunoprecipitation-mass spectrometry analysis in mitotic HEK293T cells, the researchers first identified PLK1 as a significantly enriched binding partner of RNF40. Endogenous and exogenous co-immunoprecipitation as well as GST pull-down assays further confirmed that the D4 fragment of RNF40 (222–628 aa) directly binds to the Polo-box domain (PBD) of PLK1. This suggests that RNF40 may serve as a potential recruitment scaffold for PLK1.
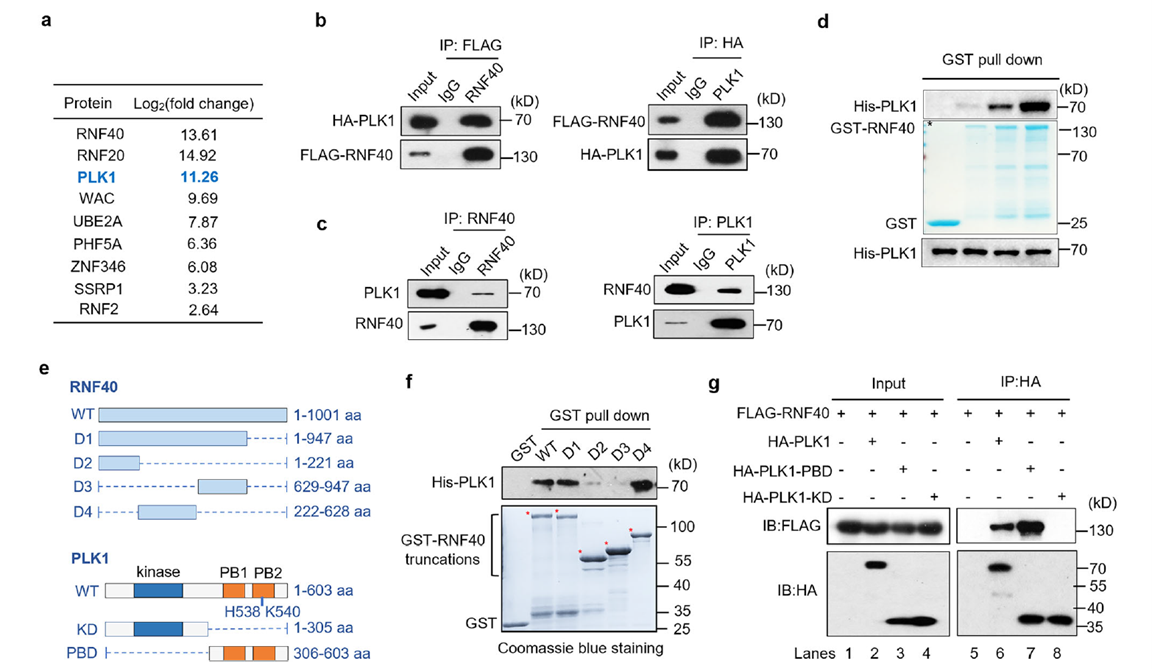
CDK1-dependent phosphorylation of RNF40 and its regulation of PLK1 binding
Sequence analysis revealed two conserved CDK1 phosphorylation sites (T529 and T557) within the RNF40 D4 fragment. Cell cycle synchronization experiments combined with phosphorylation-specific antibody detection showed that RNF40 phosphorylation begins in G2 phase and is further enhanced in M phase. In vitro kinase assays confirmed that the CDK1/cyclin complex can directly phosphorylate the GST-RNF40-D4 fragment, while the T529A/T557A double mutant (2A) completely blocks phosphorylation.
Phosphorylation directly promotes RNF40–PLK1 binding. Treatment with λ-phosphatase or CDK1 inhibitors weakens the interaction; the 2A mutant shows significantly reduced binding capacity, whereas the phosphomimetic mutant (2E) maintains strong binding. This establishes the CDK1-RNF40-PLK1 signaling cascade. At this point, the phosphorylation-regulated arm of the CDK1-RNF40-PLK1 cascade is clearly defined.
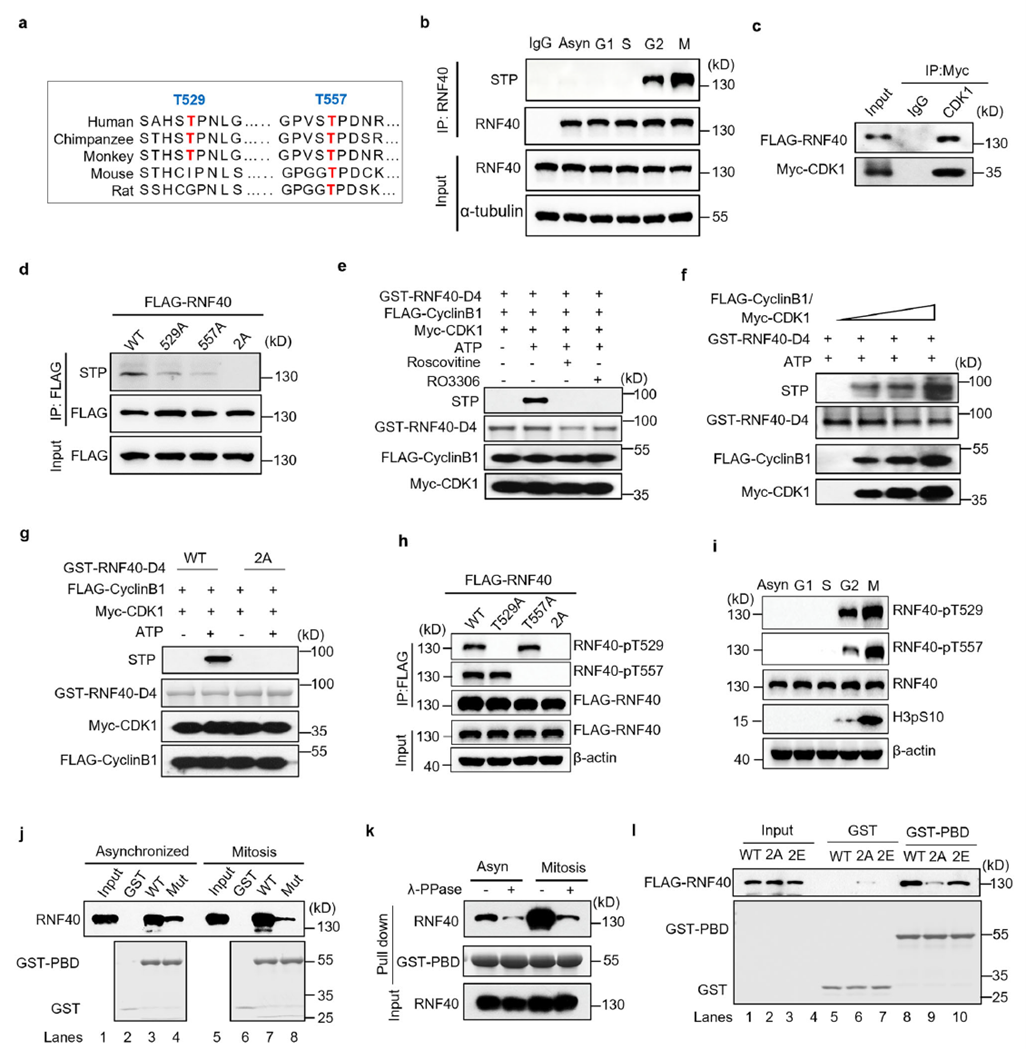
RNF40 regulates centrosome maturation and microtubule nucleation
The study confirmed that RNF40 localizes to centrosomes throughout the cell cycle, and this localization depends on the CEP192/PCNT scaffold proteins. Functional experiments showed that loss of RNF40 leads to significantly reduced PLK1 recruitment to centrosomes, which in turn impairs γ-tubulin accumulation and microtubule nucleation capacity. This functional impairment ultimately causes multipolar spindle assembly, abnormal chromosome alignment, and delayed mitotic progression. These results indicate that RNF40 acts as a key positive regulator of centrosome maturation by mediating the precise localization of PLK1.
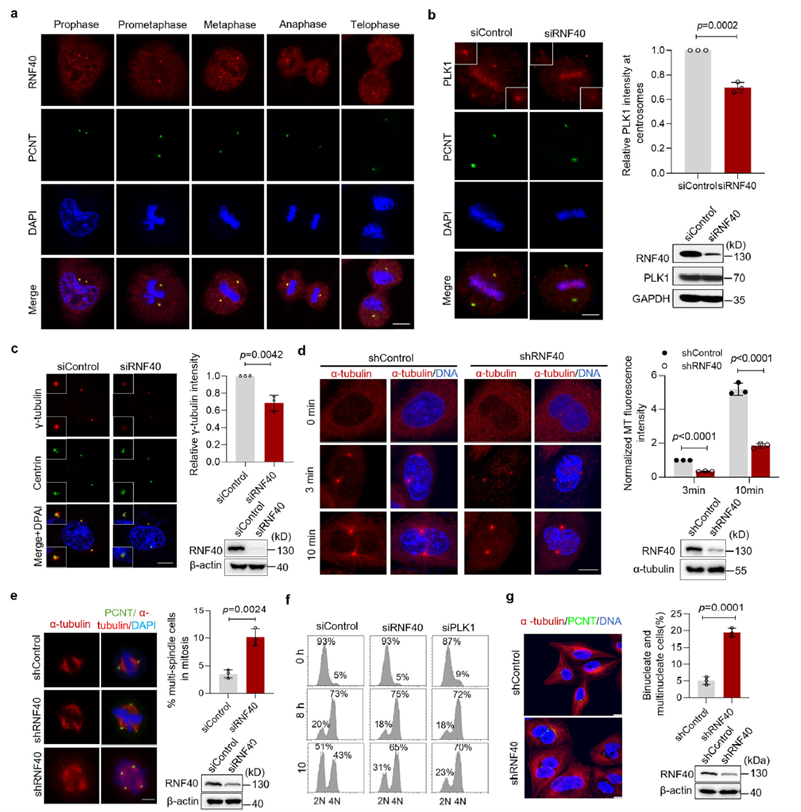
RNF40 phosphorylation is essential for maintaining chromosome stability
To verify the specific function of phosphorylation, researchers reconstituted RNF40-knockout cells with wild-type, non-phosphorylatable mutant (2A), and phosphomimetic mutant (2E). The 2A mutant failed to maintain PLK1 localization at centrosomes, leading to impaired microtubule nucleation, abnormal chromosome alignment, delayed mitotic exit, and increased multinucleated cells. In contrast, the 2E mutant fully rescued these defects. Further analysis confirmed that loss of phosphorylation (2A) directly causes abnormal chromosome numbers and genome instability. Notably, this regulatory function of RNF40 depends entirely on its phosphorylation status rather than its traditional E3 ubiquitin ligase activity.
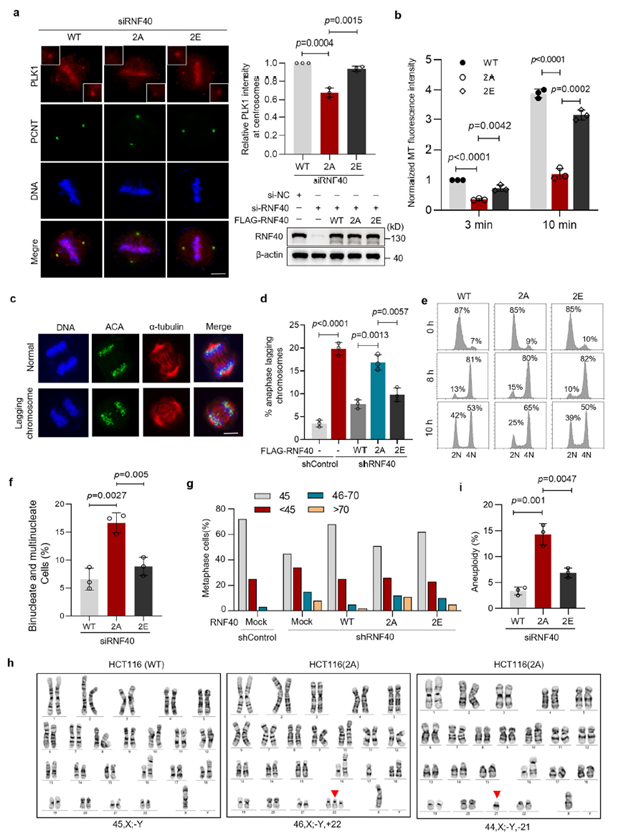
The acetylation-phosphorylation dynamic switch serves as a temporal gate for cascade activation
To explore whether RNF40 is inhibited by any modification during interphase, the researchers found that RNF40 is acetylated by PCAF during interphase (G1/S phase), and this modification is removed by HDAC1 as the cell cycle enters G2/M phase. Experiments showed a clear antagonistic relationship between RNF40 acetylation and phosphorylation: inhibiting deacetylation significantly reduced RNF40 phosphorylation levels and its binding to CDK1. This demonstrates that RNF40 undergoes a dynamic transition of “interphase acetylation lock, followed by G2/M deacetylation and phosphorylation activation.” This modification switch acts as a temporal gate for signal transduction, precisely controlling the activation timing of the CDK1-RNF40-PLK1 signaling cascade.
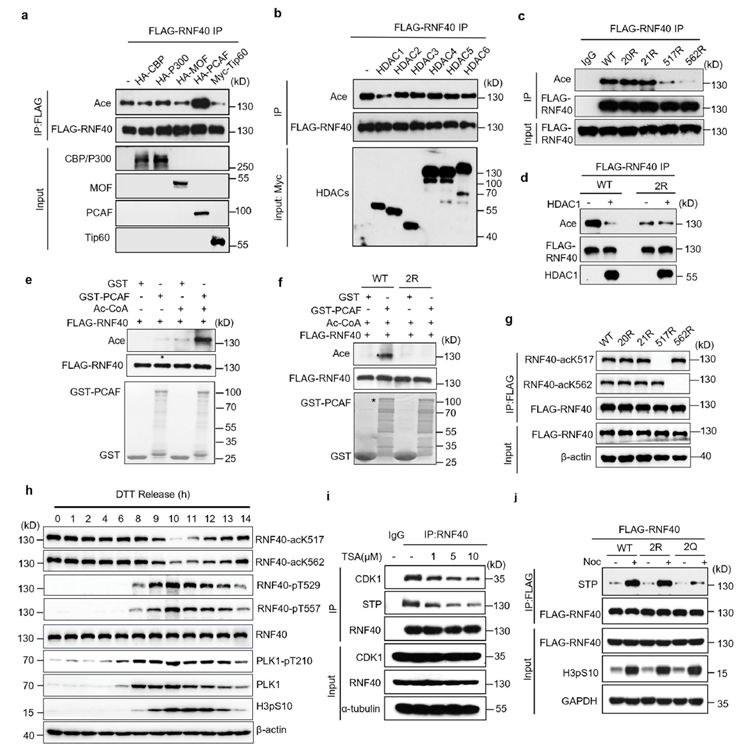
Association of RNF40 modification abnormalities with tumorigenesis and therapeutic strategies
Functional validation experiments confirmed the biological significance of the RNF40 “acetylation-phosphorylation” switch: persistent acetylation mimic (2Q) directly caused microtubule nucleation defects and increased aneuploidy, while the double-mimic mutant (2Q-2E) fully reversed the phenotype, proving that the defects stem from insufficient phosphorylation. In animal experiments, tumors expressing the acetylation-mimetic mutant (2Q) showed significantly suppressed growth, with reduced tumor volume, lower proliferation index, and increased chromosome missegregation. Clinical data further supported this finding: RNF40 is highly expressed in colorectal cancer and positively correlates with cancer stage and lymph node metastasis. In addition, specific cancer-associated mutations in RNF40 block phosphorylation, leading to centrosome dysfunction.
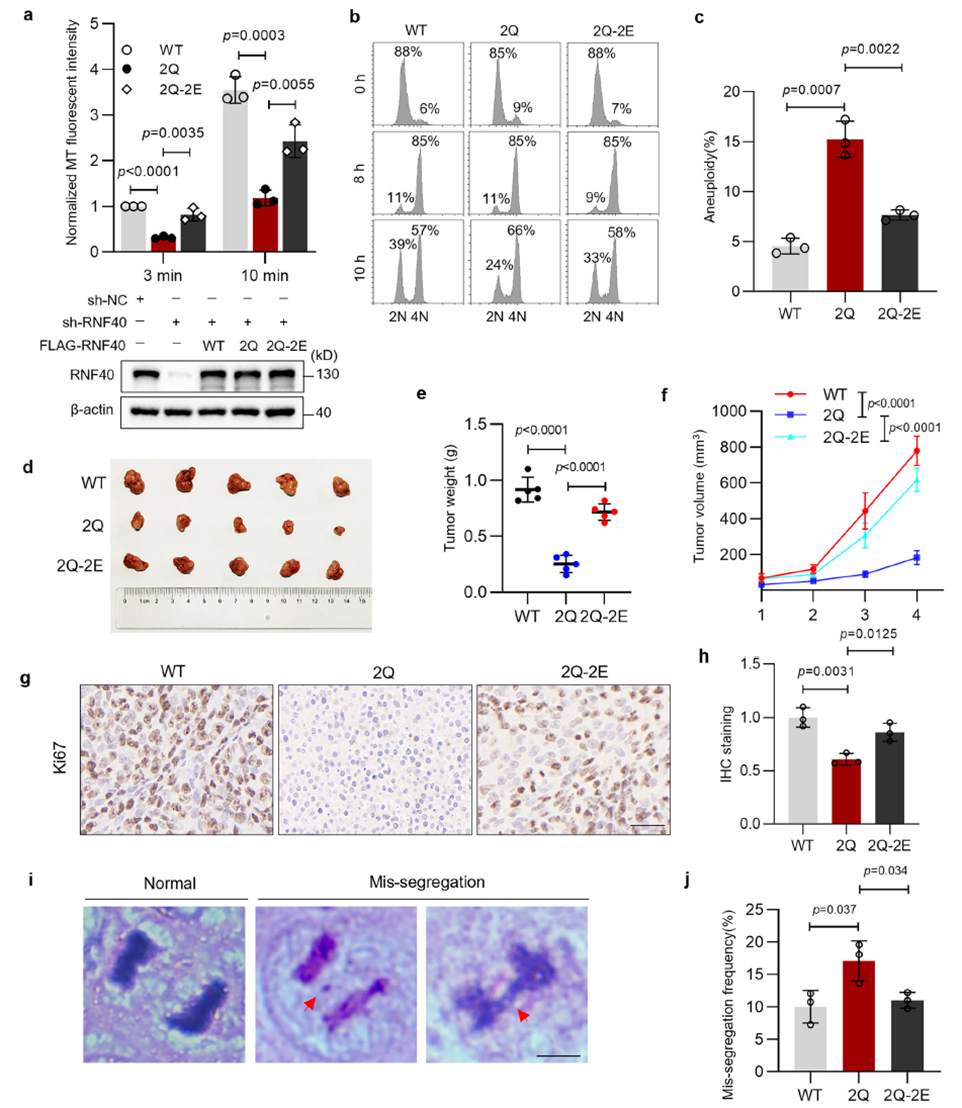
The study is the first to position RNF40 as a temporal node in centrosome maturation, revealing a precise post-translational modification switching strategy in the cell cycle. It provides a new perspective for understanding mitotic fidelity and tumorigenesis, and opens a new window for developing anti-cancer strategies targeting centrosome-spindle abnormalities.
AtaGenix Technical Support
AtaGenix custom-developed highly specific RNF40 phosphorylation antibodies (pT529, pT557) and acetylation antibodies (acK517, acK562), enabling the research team to clearly capture the dynamic changes in RNF40 modification states across the cell cycle. These antibodies provided critical evidence for establishing the temporal window of the CDK1-RNF40-PLK1 signaling cascade and laid a solid experimental foundation for the subsequent development of anti-cancer strategies targeting post-translational modifications.
AtaGenix specializes in custom post-translational modification antibody development with a focus on high-specificity antibodies
Successfully supporting multiple high-impact studies in cell cycle and tumor mechanisms
Contact us now to customize your phosphorylation/acetylation antibodies
Contact Us
+86-27-65523339
info@atagenix.com
Building C, R & D Building, No. 666, Shendun 4th Road, Donghu New Technology Development Zone, Wuhan
